Decision making is not limited to animals like humans or birds. Bacteria also make decisions with intricate precision. Imagine being so tiny that you are literally moved by water molecules bumping into you. This is what bacteria encounter perpetually. Now, imagine having no eyes, no ears, no sense of touch, no taste or nose. How would you know what or who was around you? How would you find food now as compared to where you were a short time ago? This is where being able to sense important things like a food source is critical. Bacteria have this on their “mind” all the time. Depending on the size of a bacterium’s genome, these tiny organisms have the ability to sense hundreds to thousands of internal and external signals like carbon sources, nitrogen sources, and pH changes. If these bacteria are motile (able to move around), they can compare how conditions are for them now against how they were a few seconds ago. That’s right, bacteria have a memory albeit short. If conditions are better, they can continue to move in a forward direction. If conditions are worse compared to a few seconds earlier, they can change direction and continue searching for better conditions in their environment to generate energy. But, how do they decide?
I will focus on a lesser known bacterium as my example since I have the most knowledge about it. Azospirillum brasilense is found in the soil around the world and interacts with the roots of cereal plants like corn and wheat. A. brasilense is almost always (except when attached to plant roots) motile and searching for the best niche to provide energy for the cell. This bacterium can “make” its own usable form of nitrogen from nitrogen gas in the air through a process known as nitrogen fixation. This costs the cell a lot of energy so they are searching for nitrogen sources as well as the necessary carbon sources for life. The microscopic world can be cut throat. Having the ability to sense a greater variety of food compounds could mean the difference between being the predominant species in town or being on the fringe.
Back to the question about how these cells decide which direction to travel. One way is through a dedicated group of proteins that regulate how often the cell switches direction. This group of proteins control chemotaxis, the movement of a cell in response to chemicals within their environment. The number of chemotaxis genes varies depending on the complexity of metabolism for a bacterium. The champion at the moment is 129 fromPseudomonas syringae pv. oryzae str. 1_6. The proteins that actually sense the chemical signal are called methyl-accepting chemotaxis proteins (MCPs) or chemoreceptors. Azosprillum brasilense has 48 MCPs within its genome. This does not mean, however, that A. brasilense cells can only sense 48 different chemicals. Most, but not all, of these MCPs don’t interact with the chemicals themselves but sense the changes in the amount of energy the cell has within the environment they reside. If things are good, the MCPs are inactive. However, if energy levels are lower than they were a few seconds before, the MCPs become active and begin the signal to change direction. And these MCPs are VERY sensitive to changes. For example, if the A. brasilense cells are swimming in a liquid medium with 1,000 molecules of sugar, they will detect changes of addition or removal of a few sugar molecules in the medium. Now, move these cells immediately into a medium with 1,000,000,000,0000 sugar molecules and they still will be able to detect removal or addition of a few sugar molecules. This is called adaptation and allows the cells to remain sensitive no matter the concentrations of compounds they encounter.
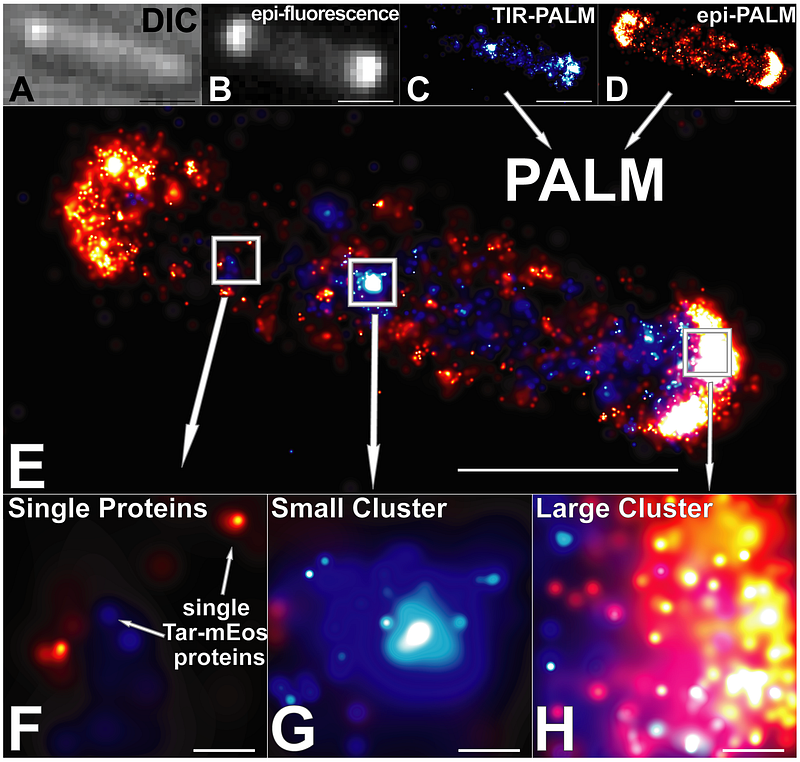
MCPs by themselves would be useless if they did not interact with some other cellular machinery. Lucky for bacteria, the MCPs are only the first of many specialized proteins for regulating direction of travel. If a chemical binds to an MCP outside the cell, how does the inside of the cell get the message? When chemicals bind to an MCP or stop binding to it, it slightly changes the structure of the MCP. It is thought association or dissociation of chemicals to MCPs causes a rotation like a slight turn of a door knob. Depending on if the MCPs are rotated or not changes the activity of another enzyme that interacts with MCPs, a protein called CheA (pronounced, ‘key A’). When CheA is active, it converts two other proteins into an active form, CheB and CheY. Confused yet? When CheY is in its active form, it interacts with the base of the flagellumand causes the flagellum to switch direction of rotation and causes the bacterium to change the direction it is moving. The take home message for the mechanism is this: when the MCPs are not interacting with chemicals (nutrients), CheA and CheY are active, and the flagellum switches rotation to make the cell go in a different direction. Hopefully for the cell, the new direction it is traveling will have more nutrients that will interact with the MCPs and block more changes in direction by CheA/CheY activity.
You might ask, if chemical compounds are always bound to the MCPs, what if the cell begins going to passed the best environment and needs to turn around? Good question. That is where the other protein activated by CheA comes in, CheB. What makes this system unique is the ability to adapt to current conditions (nutrient chemical levels) so the cell can respond to new information. A set of protein enzymes act upon part of the MCPs that can cause a change in how rotated (think door knob again) the MCP is. If the MCP is always interacting with a nutrient, a protein called CheR changes the MCP structure and causes rotation back towards the non-interacting form of the MCP. When CheA is active, it can activate CheB which reverses the changes caused by CheR. Ultimately, the whole system remains sensitive to new information over a wide range of nutrient concentrations (a 1000-fold range).
This whole system and its parts and regulation took several years for me to understand. I’m sure I did not do it justice, but hopefully you can get a small glimpse into the truth about “simple” bacteria.

One of the most prevalent ways a bacterium decides this is by using a two component system, or TCS. TCSs are relatively simple compared to chemotaxis. As you would suspect from the name, TCSs are pathways consisting of only two protein members, the sensor histidine kinase and the response regulator. Histidine kinases are a major protein family in bacteria because they are able to sense many different factors in the bacterium’s environment including nutrients, toxins, fellow bacteria, etc. In case you are wondering, chemotaxis is a modified form of a TCS in which the histidine kinase CheA is regulated by the activity of a separate protein, the methyl-accepting chemotaxis protein.
What if you are a bacterium and you have been using a certain type of carbon source to generate energy and suddenly that carbon source isn’t as prevalent? In this case, you would want to shut down the enzyme factories that were converting the previous food source into energy and begin preparing new enzyme factories to convert other food sources into energy as you prepare for starvation. If these conditions persist, you might want to decide to hibernate in the form of a spore or cyst until conditions around you improve. Or, if other food sources are sensed in the environment, any special enzymes that would be needed to convert them into energy would need to be synthesized from their respective genes. All of these scenarios are controlled by TCSs. The conditions are used as input for the cell to decide the best strategy to survive and thrive. Histidine kinase activation leads to a hand-off event from the kinase to the response regulator of a molecule which acts as a green light for the response regulator to proceed with its job. This job may be to turn on gene expression to produce proteins needed in the cell. The response regulator’s job may be to shut down gene expression for proteins no longer needed by the cell. It is a carefully orchestrated balancing act evolved over millions of years to make sure only the proteins/enzymes needed by the cell at a given time are present assuring highly valuable energy molecules are not wasted.
Getting the message
Second messengers are common from bacteria to humans. The major second messenger we all learn about in biochemistry class is cyclic AMP (cAMP). However, bacteria use several nucleotides as second messengers. Many are used as determinants in the decision making process, but one of the most recently discovered (and personal favorite) is cyclic-di-GMP, or c-di-GMP.
Bacteria are constantly processing signals both inside and outside their cell membranes. Hard to believe that one of the most abundant response molecules was only discovered in the late 1980s while researching how a certain species, Acetobacter xylinum now known as Gluconacetobacter xylinus, produced cellulose. Almost by accident, the Benziman lab discovered the enzyme responsible for cellulose production (cellulose synthase) was regulated by a nucleotide, later found to be c-di-GMP. Since that discovery, c-di-GMP has become a hot topic among microbiologists and immunologists due to the decisions bacteria make as the level of c-di-GMP increases within the cell. As I learned it, the concentration of c-di-GMP had predictable outcomes on the decisions of bacteria: high levels leads to loss of motility, increase in biofilm formation, changes in cell morphology, and increase in cell-cell communication. When low levels of c-di-GMP are present, the cell decides to move around (motility), become resistant to heavy metals, and, most importantly, becomes virulent. For example, Vibrio cholerae, the bad guy responsible for cholera, only decides to move around and produce cholera toxin when c-di-GMP levels are low in the cell. If levels increase, V. choleraewill produce biofilms via extracellular polysaccharide (EPS) production.
You might be asking yourself what controls the c-di-GMP levels of a bacterial cell. The initial discovery in the Benziman lab also found the enzymes/proteins that were responsible for making and breaking the second messenger. The long (and short) names are; for making c-di-GMP from 2 GTP molecules, diguanylate cyclases (DGCs aka GGDEF proteins) and degradation by phosphodiesterases (PDEs aka EAL proteins). GGDEF and EAL proteins are so called due to important amino acids necessary for their functions, GGDEF is glycine, glycine, aspartate, glutamate, phenylalanine, and EAL is glutamate, alanine, leucine. These enzymatic activities are usually controlled by regulatory protein domains common in bacteria (and humans). Signals from the environment (internal or external) can trigger changes in enzyme activity of GGDEFs and EALs thus changing the cellular concentration of c-di-GMP. This mechanism is well understood after 30 years of research. However, what happens next is still essentially unknown.
Cyclic-di-GMP levels rise within a bacterial cell. Now what? It was known early on that c-di-GMP itself could then interact with GGDEFs to inhibit activity. But what other proteins interact with c-di-GMP and help these bacteria decide to make major lifestyle changes? It wasn’t until 2006 that bioinformaticians predicted c-di-GMP binding to a protein, or protein domain. PilZ, an obscure protein of unknown function but necessary for Type IV pilimotility, was hypothesized to bind c-di-GMP. By the end of 2007, this prediction was verified and PilZ domain proteins were the first shown linking c-di-GMP to downstream proteins in pathways, or circus rings.
Transitioning from a free swimming/moving cell to life in a biofilm community is a major lifestyle change for bacteria. This decision takes commitment which is initiated by a small molecule.
Mystery of a mysterious kind
The decisions bacteria make weigh heavily on the amount of this molecule found within the cell. However, how a bacterial cell knows how much c-di-GMP there is ultimately remains a mystery. The focus of early research involved finding what regulated the synthesis and degradation of c-di-GMP. Thus, the majority of publications in print focus on the enzymes that perform these functions, GGDEFs and EALs. Also, by deleting certain GGDEFs or EALs from a bacterium, scientists were able to determine what effect this change would have on the cell’s decision making and lifestyle. Most research was performed in medically relevant species like Vibrio and Pseudomonas. This short-sighted focus has led to a distinct role of c-di-GMP in the cell that may not be absolute. I digress…
A cell produces c-di-GMP in response to some environmental signal. Now what?
Great question; one that is still not answered. Various proteins have been shown to interact with or bind c-di-GMP including the PilZ domain. The list of c-di-GMP effectors has grown slightly over the past few years to include examples of transcriptional regulators in both Vibrio and Pseudomonas (VpsT and FleQ, respectively). Transcriptional regulators are proteins that help carry out the decisions made by a cell by regulating gene transcription. However, these are only two examples from two bacteria. What about the vast number of other bacteria out there? How do they “see” c-di-GMP? Cyclic-di-GMP is a vital component to bacterial decision making even though our knowledge of how it is seen by a cell is a huge unknown.
My hypotheses and speculations (with some evidence)
In the last chapter of my dissertation (under embargo), I investigated what other protein domains could potentially bind c-di-GMP bioinformatically. Using my methods, I could predict proteins in the nonredundant database that could potentially bind c-di-GMP. One group of proteins I found were those already shown to bind other nucleotides like ATP. I was able to test my method against a publication that biochemistry and proteomics to identify c-di-GMP binding proteins from Pseudomonas. This crude “chemical proteomic” approach identified around 200 potential binding proteins of which the method I created also found several of the same proteins without the exhaustive time and effort of “wet bench” experiments.
This is not a post about how good I am at science. This is a post about using new and different methods to answer questions within science not unlike the investigation that identified the PilZ domain as the c-di-GMP binding protein in the first place. Unfortunately, my time in the lab was over before I could test my hypotheses, but my curiosity and passion live on. I will say that I predict receiver domains are very common c-di-GMP binding effectors that will be the next major discovery in this elusive mystery of how cells use c-di-GMP to make decisions.
It takes a village
The marvels of single celled organisms is that they are able to integrate all kinds of stimuli and make one grand decision that affects how they proceed. Bacteria do in one cell what we as humans do with billions. However, do bacteria contain the ability to think as a group or community?
The answer is absolutely. It is called quorum sensing. The pioneer for this research is Bonnie Bassler from Princeton University. Listening to her tell her story of the curiosity she felt when observing how and why a certain group of bacteria emitted light, or bio-luminescence is great. (Watch here). Through her investigation with a insignificant bacterium, Vibrio harveyi, she opened up a whole new field of microbiology.
Many bacteria synthesize signaling molecules that serve as messages to other bacteria saying, “I am here”. Since bacteria don’t have senses that we are familiar with like sight and hearing, these signaling molecules tell other bacteria who is around. When there aren’t a lot of bacteria sending out the signal, no big decisions are made. However, when enough bacteria are around to tell all other village members the approximate population, all village members act together to make a committed decision. In the case of V. harveyiit is the production of a light emitting molecule, but for other bacterial species it may be activation of pathogenicity. From the perspective of the bacterium, you don’t want to decide alone to make a big commitment like invading another organism. By taking a bacterial census through quorum sensing, these bacteria make a educated decision only when their population is high enough to make an impact. For some species, this critical number may be less than ten. However, in some cases, the population needs to be in the millions.
I think bacteria can teach us a very important lesson via quorum sensing: don’t go it alone. It takes a village.
